Science | 清華大學和南京農大揭示植物抗病基因PGIPs作用機制


2024年2月15日,Science在線發表了由清華大學和南京農業大學合作的研究內容“A plant mechanism of hijacking pathogen virulence factors to trigger innate immunity”。本研究表明,植物在響應病原體入侵過程中,參與抑制病毒聚半乳糖醛酸酶活性的基因PvPGIP2與FpPG(聚半乳糖醛酸酶)的相互作用主要包括兩個方面:
1.抑制FpPG酶的活性,促進FpPG水解PGA產生更多的長鏈低聚半乳糖醛酸酯,同時減少短鏈低聚半乳糖醛酸酯的產生,進一步可以促進PTI的產生;
2.PvPGIP2結合到上FpPG上之后產生一個底物結合位點,其底物結合活性增強,底物偏好改變。基于結構工程可以將最初缺乏FpPG結合活性的推定PGIP轉化為有效的FpPG相互作用蛋白。
具體的研究內容如下:
研究背景:
植物細胞壁由纖維素、半纖維素和果膠組成,是抵御病原體侵襲的主要和關鍵屏障。病原體分泌各種細胞壁降解酶(CWDEs),如聚半乳糖醛酸酶,以解聚細胞壁多糖,并隨后破壞細胞壁的完整性。聚半乳糖醛酸酶由各種病原體產生,它們在發病機制中的作用已經得到了很好的證實。病原性和某些植物源性聚半乳糖醛酸酶屬于內糖苷水解酶28家族(內聚半乳糖醛酸酶)。這些酶非特異性地水解細胞壁中α-1,4連接的聚半乳糖醛酸(PGAs,果膠的去酯化高半乳糖醛酸)的糖苷鍵。為了抵消CWDE的酶活性,植物表達了一系列CWDE抑制蛋白,包括PGIPs。此外,植物已經進化出細胞膜上的模式識別受體(PRRs)來識別CWDEs和/或它們釋放的細胞壁片段作為損傷相關的分子模式,從而誘導模式觸發免疫(PTI)。例如,擬南芥PRRs壁相關激酶(WAKs)識別聚合度為10至15度的長鏈低聚半乳糖醛酸酯(OG10-15),作為PTI激發子。因此,病原體多半乳糖醛酸酶在破壞植物細胞壁結構以增強毒力和向植物發出信號以觸發防御反應方面具有雙重作用。
PGIPs是富含亮氨酸重復序列(LRR)蛋白家族的成員,廣泛存在于單子葉植物和雙子葉植物中。作為多半乳糖醛酸酶的特異性和高親和力受體,最具特征的PGIP, Phaseolus vulgaris isoform 2 (PvPGIP2),可以識別幾種真菌多半乳糖醛酸酶,包括葉根鐮刀菌多半乳糖醛酸酶(FpPG)。CWDE抑制蛋白以一種保守的機制結合CWDE的活性位點。PvPGIP2被提出結合活性位點裂縫使FpPG和炭疽菌多半乳糖醛酸酶1 (CluPG1)失活。除了限制聚半乳糖醛酸酶的破壞潛力外,PGIP-PG相互作用還促進了體外和表達PGIP-PG嵌合體的轉基因擬南芥中誘導活性低聚半乳糖醛酸酯的積累。炭黑菌多半乳糖醛酸酶1 (ClPG1)的酶活性對于產生煙草防御反應的激發子至關重要。PGIPs在提高抗病性方面的有效性已在多種作物中得到證實,包括小麥和水稻。然而PGIPs的作用機制仍不清楚。
研究結果:
一、PvPGIP2改變FpPG催化的低聚半乳糖醛酸酯長鏈和短鏈比例
1.確定PvPGIP2與FpPG之間存在相互作用
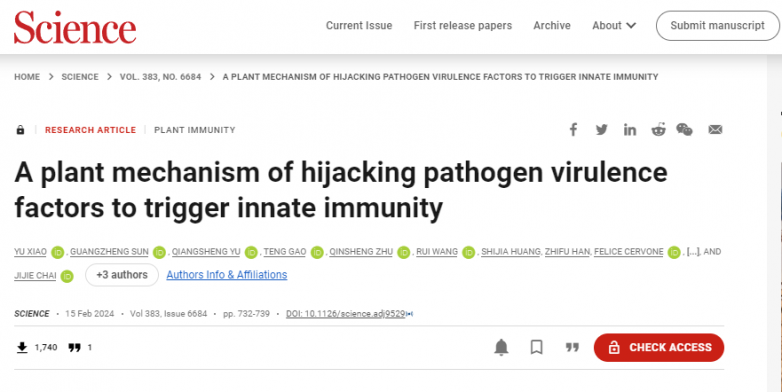
為了重建PvPGIP2和FpPG之間的相互作用,作者在昆蟲細胞中分別表達了這兩種蛋白。凝膠過濾和分析性超離心實驗表明,這兩個蛋白在pH 5.0下形成了穩定的復合物,分子比為1:1(圖1A和B)。等溫滴定量熱法(ITC)進一步證實了PvPGIP2-FpPG的相互作用,FpPG與PvPGIP2的解離常數(Kd)為15 nM(圖1C)。堿性條件(pH 8.0)完全破壞了PvPGIP2FpPG相互作用,這表明PvPGIP2可能在植物免疫應答的早期階段發揮作用。鹽濃度的增加略微降低了它們的結合親和力。
2.確定PvPGIP2與FpPG之間的相互作用確實會影響PTI的程度
先前描述的高性能陰離子交換色譜脈沖安培檢測(HPAEC-PAD)測定用于研究以PGA為底物,在PvPGIP2存在或不存在的情況下,FpPG催化的低聚半乳糖醛酸產物的大小分布。與之前的數據一致,添加PvPGIP2降低了FpPG催化的PGA水解,表現為生成更多的OG>9和更少的OG<9(圖1D)。酶學特征數據支持PvPGIP2抑制FpPG酶活性。
OG10-15激活廣泛的PTI防御反應,因此作者首先用FpPG測試了PGA消化產物的PTI誘導活性。接種大豆疫霉P6497前,用dH2O、OG10-15、FpPG+PGA或FpPG- PvPGIP2 +PGA處理大豆黃化下胚軸24小時。接種P649748小時后,用FpPG-PvPGIP2+PGA或OG10-15處理的黃化下胚軸沒有明顯的疾病發展癥狀。相比之下,用dH2O、PGA或FpPG+PGA預處理的黃化下胚軸可以觀察到明顯的疾病癥狀(圖1E)。與此一致,FpPG-PvPGIP2+PGA或OG10-15處理的黃化下胚軸中P6497的生物量顯著降低(圖1F)。FpPGPvPGIP2+PGA或純OG10-15比FpPG+PGA或單獨使用PGA處理中活性氧(ROS)的產生(圖1G)和大豆PTI標記基因細胞色素P450 93家族(GmCYP93)的表達(圖1G)程度更大。FpPG-PvPGIP2+PGA更強的PTI誘導活性進一步通過分析擬南芥中胼膜沉積(圖1I和J)和PTI相關基因WRKY DNA結合蛋白40 (WRKY40)、植物恢復素缺陷3 (PAD3)、鈣調蛋白樣41 (CML41)和AtPGIP1的表達得到證實(圖1K)。
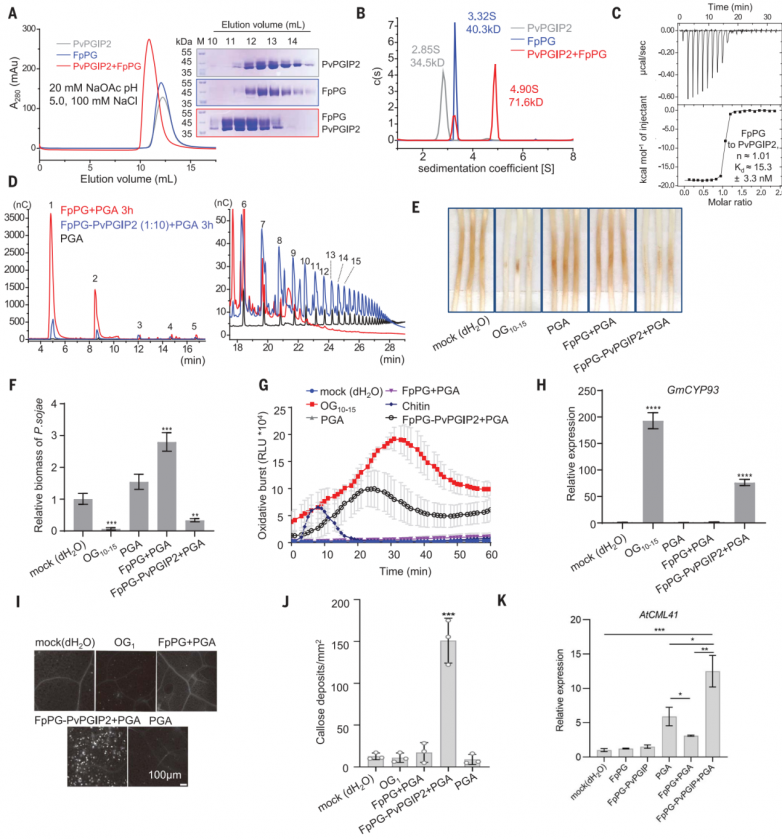
圖1.PvPGIP2-FpPG產生誘導活性低聚半乳糖醛酸酯
3.影響PTI程度的主要是短低聚半乳糖醛酸酯
FpPG+PGA混合物含有大量短OGs(圖2A),已被證明可以抑制小麥的防御反應。為了測試合成的短低聚半乳糖醛酸酯是否影響植物免疫,作者對OG2-7,OG5-7或OG3對大豆中不同病原體相關分子模式(PAMPs)和OG10-15激活的PTI反應的影響進行測試。OG2-7、OG5-7或OG3預孵育24小時,顯著抑制了大豆中flg22、幾丁質、疫霉1 (INF1)或OG10-15誘導的氧化爆發產生(圖2B)和防御基因GmCYP93、致病相關蛋白2 (GmPR2)和1-氨基環丙烷-1-羧酸合成酶2 (GmACS2)(圖2C)的表達。與OG10-15相比,OG2-7、OG5-7或OG3在大豆中沒有引發ROS的產生(圖2B)。這些結果表明,短的低聚半乳糖醛酸酯可以作為PTI信號傳導的一般抑制劑,盡管其潛在機制尚不清楚。FpPGPvPGIP2+PGA混合物也含有短的低聚半乳糖醛酸酯(OG<9),這一觀察結果可以解釋為什么FpPG-PvPGIP2+PGA引發的免疫反應比OG10-15弱得多(圖1,E至H)。
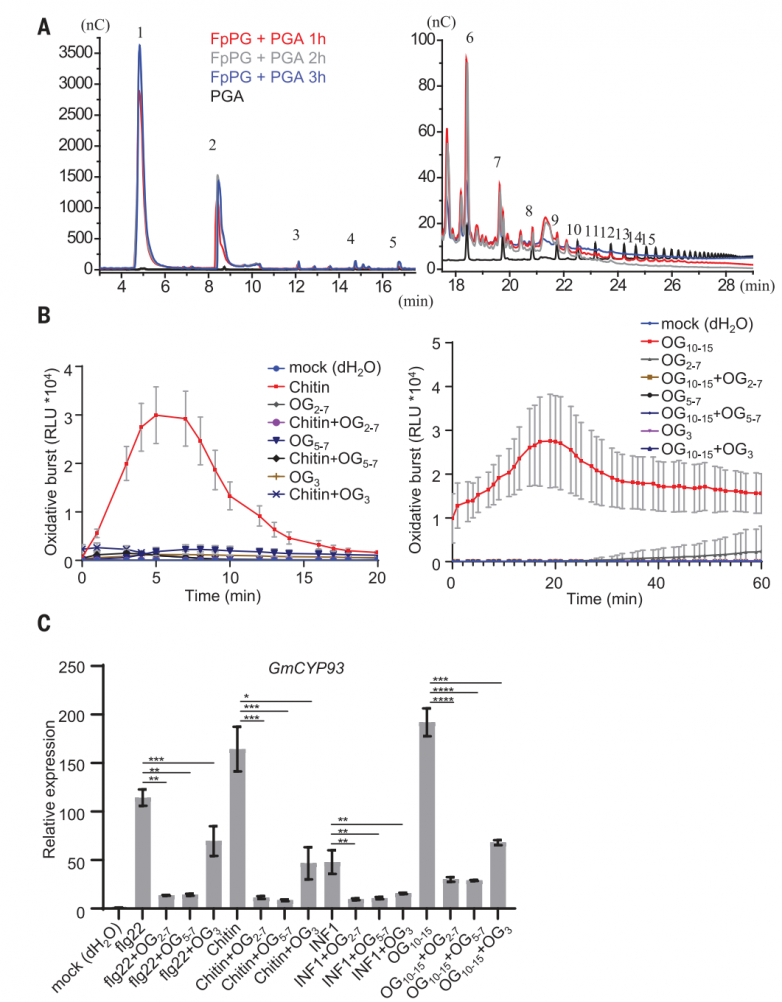
圖2.FpPG產生免疫抑制劑低聚半乳糖醛酸酯
二、PvPGIP2N274D-FpPG的整體結構及識別機制
最初試圖結晶PvPGIP2-FpPG復合物的努力失敗了,可能是因為糖基化會干擾分泌蛋白的結晶。PvPGIP2-FpPG突變復合蛋白的一個預測糖基化位點PvPGIP2 Asn274被Asp取代(PvPGIP2N274D)成功結晶。用分子置換法求解晶體結構,并將其細化為1.93 ?。
晶體中,PvPGIP2N274D與FpPG形成1:1的配合物,埋面為1087.1 ?2(圖3A)。FpPG折疊成右側平行的β螺旋,這是典型的果膠甲基酯酶、水解酶和裂解酶的結構。β螺旋包含10個完整的三股或四股螺旋,它們具有不同的大小和構象,從核心區域伸出。預測的FpPG D191、D212和D213的活性位點殘基位于第3圈(T3)環上。PvPGIP2N274D的結構包含10個串聯重復的LRRs(圖3A),與其他植物中含有LRR的蛋白一樣,折疊成右手超螺旋。FpPG結合的PvPGIP2N274D的構象與游離的PvPGIP2幾乎相同,這表明與FpPG的相互作用不會引起PvPGIP2N274D的構象變化。1匝(T1)環區和c端側平行的β鏈(PB) 2a在很大程度上介導了FpPG與PvPGIP2N274D的N端內表面的相互作用(圖3A)。相互作用使PvPGIP2N274D的C端靠近FpPG的β螺旋2至5的長T3環,導致FpPG頂部活性位點關閉,并在PvPGIP2N274D-FpPG復合物中形成活性位點隧道(圖3B)。
PB2a垂直包裹在PvPGIP2N274D的N端5條β鏈上。PvPGIP2N274D的四個芳香殘基與FpPG的接觸主導了該區域周圍的相互作用(圖3C)。來自PvPGIP2N274D第一個LRR的Asp56通過與FpPG Asp250和Asn279形成分叉氫鍵進一步加強了相互作用(圖3C)。除了PB2介導的相互作用外,三個T1環進一步促進了PvPGIP2N274D-FpPG相互作用(圖3D)。來自β-螺旋9的FpPG Gly314與PvPGIP2 Val152疏水接觸,而FpPG的Asn313、Gly343、Ser344和Lys316與PvPGIP2 Tyr105相互作用(圖3C)。PvPGIP2 Thr177和Phe201在β-螺旋7/8的T1中心區域進行了組裝(圖3D)。FpPG的Ala274位于該區域但難以接近(圖3D)。在FvPG中,FpPG Ala274突變為體積較大的Thr274,從而消除了PvPGIP2-FpPG相互作用。相反,T274A足以賦予FvPG與PvPGIP2相互作用的能力。Gln224對于PvPGIP2與FpPG的相互作用很重要,它與FpPG Asn304和FpPG Ser272分別在-β螺旋9和β-螺旋8中建立了雙齒氫鍵(圖3D)。FpPG Asn304也與PvPGIP2 Phe269形成范德華接觸(圖3D)。與PvPGIP2相互作用至關重要的FpPG殘基在其他PvPGIP2結合的聚半乳糖醛酸酶中大部分是保守的。相比之下,PvPGIP2中關鍵的FpPG相互作用殘基在不同植物的PGIPs中保守性較差,特別是在非豆科植物中。
接下來,作者對PvPGIP2和FpPG進行了氨基酸替換,并使用ITC評估了這些突變對PvPGIP2-FpPG相互作用的影響。Y105E預計會破壞與FpPG殘基簇的相互作用(圖3C),將PvPGIP2與FpPG結合的結合親和力降低了900倍。與FpPG T1中心區域相互作用的PvPGIP2 Phe201突變(圖3D)類似地損害了PvPGIP2-FpPG相互作用(圖3C)。PvPGIP2 V152E破壞了與FpPG β-helix9的相互作用,以Kd ~2.57 mM結合FpPG,效率比野生型PvPGIP2低約170倍。PvPGIP2 β8(圖3D)下的FpPG Thr275突變使FpPG與Tyr的PvPGIP2結合親和力降低了2000多倍。HPAEC-PAD分析表明,這些PvPGIP2-FpPG突變蛋白在水解PGA時產生的低聚半乳糖醛酸譜與單獨的FpPG相似。
擬南芥多半乳糖醛酸酶側根(PGLR)的多半乳糖醛酸酶活性已被證實。PGLR的C端部分,對應于FpPG的PvPGIP2接觸表面,被一個額外的C端的α螺旋完全阻斷,這在其他一些假定的擬南芥多半乳糖醛酸酶中是保守的。因此,ITC沒有發現PGLR和AtPGIP1之間的相互作用。這些數據進一步支持了FpPG C端與PvPGIP2相互作用的相關性。然而單個PGIP可以采用不同的PG抑制機制。
圖3.PvPGIP2不阻斷FpPG的活性位點
三、PvPGIP2促進免疫活性低聚半乳糖醛酸酯產生的機制
阻斷活性位點被認為是PGIP抑制聚半乳糖醛酸酶的機制。然而,PvPGIP2不結合FpPG的活性位點(圖3B)。已知PvPGIP1和PvPGIP2具有PGA結合活性。ITC實驗表明PGA,而不是其他多糖,在pH 5.0時結合PvPGIP2。PGA的結合活性,在其他物種的PGIPs中是保守的,在pH 8.0時完全被消除,但與Ca2+無關。OG10-15和PvPGIP2之間沒有相互作用,可能是因為在PGA中發現的21/31螺旋結構不能在OG10-15中有效形成,但對PvPGIP2識別PGA至關重要。
PGA結合活性表明PvPGIP2與FpPG形成復合物時,可以增強FpPG對底物的結合親和力。為了量化FpPG和PvPGIP2-FpPG的底物結合親和力,使用PGA水解能力嚴重受損的FpPGH234K變體來最大限度地減少底物降解。由于分子量未知,PGA結合FpPGH234K無法通過ITC準確定量。因此選擇了市售的OG10-15作為PGA的代理。雖然OG10-15與PvPGIP2之間沒有相互作用,但PvPGIP2-FpPGH234K顯示出比FpPGH234K (圖4A)更高的OG10-15結合親和力(圖4A)。相比之下,低聚半乳糖醛酸酯(OG5-7)與PvPGIP2-FpPGH234K的親和力要弱得多。正如預期的那樣,在相同的條件下,OG5-7和FpPGH234K之間沒有相互作用。這些結果表明,酶活性較低的FpPG變體可能更有效地產生較長的低半乳糖醛酸酯。事實上,FpPGH234K或FpPGH234A仍然保留了殘余的PGA水解活性,比野生型FpPG產生更多的長鏈低聚半乳糖醛酸酯(圖4B)。FpPGH234K或FpPGH234A催化活性聚半乳糖醛酸酯的產生在大豆(圖4C至F)和擬南芥(圖4G至I)中得到了證實。與FpPGH234K或FpPGH234A相比,PvPGIP2-FpPGH234K或PvPGIP2-FpPGH234A沒有PGA水解活性(圖4B)。
FpPGH234K或FpPGH234A催化產物的免疫誘導活性低于PvPGIP2-FpPG降解產物。這一結果表明,除了抑制外,其他機制也可能參與促進PvPGIP2-FpPG產生免疫活性低聚半乳糖醛酸酯。PvPGIP2表面上由Arg183、Arg206、Lys230和Arg252組成的帶正電簇是PGA結合所必需的。該簇在不同植物物種的PGIPs中是保守的,但它不參與與FpPG的相互作用(圖4J)。該簇位于FpPG-PvPGIP2活性位點通道的連續體中,表明它是底物結合酶復合物活性位點的一部分(圖4J)。四殘基丙氨酸(PvPGIP24A)同時突變極大地削弱了FpPGH234K-PvPGIP2的OG10 -15結合親和力(圖4A)。相比之下,OG5-7與FpPGH234K-PvPGIP2的結合被完全消除。HPAEC-PAD分析表明,FpPG- PvPGIP24A或FpPGH234K-PvPGIP24A產生的低聚半乳糖醛酸酯的大小分布與FpPG或FpPGH234K催化的低聚半乳糖醛酸酯相似(圖4B),這表明PvPGIP2突變消除了對FpPG的抑制作用。綜上所述,這些結果表明PvPGIP2在PvPGIP2- FpPG復合物中創建了一個底物結合位點,該位點優先結合OG10-15而不是OG5-7。
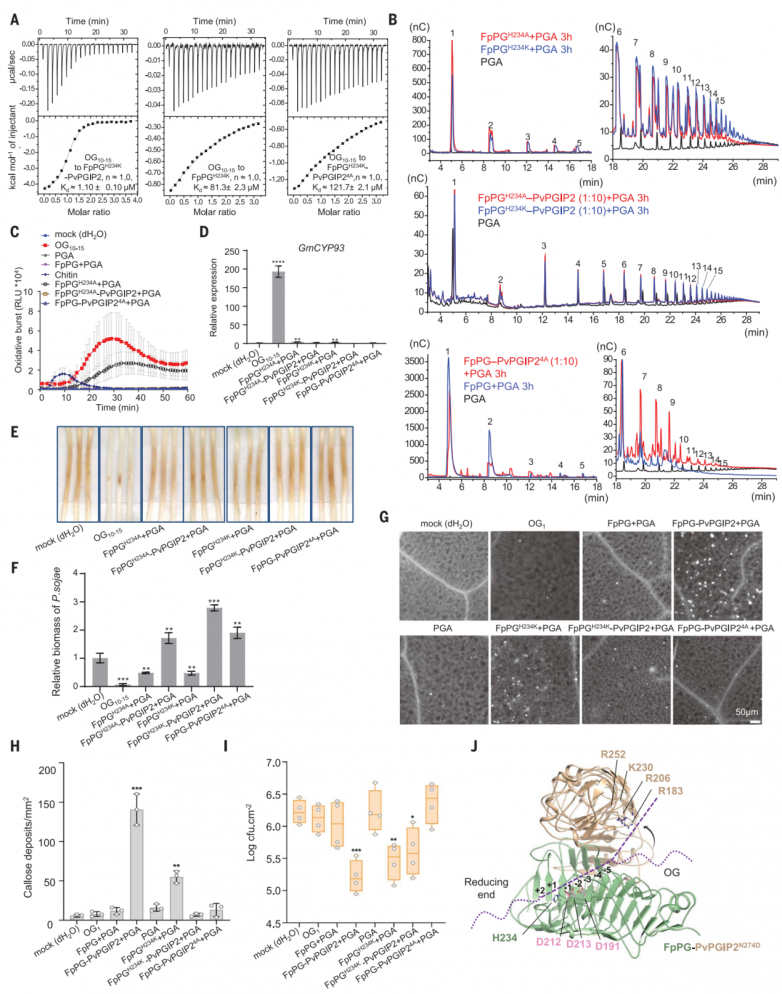
圖4.pvpgip2促進免疫活性低聚半乳糖醛酸酯產生的機制
邁維代謝現推出修飾蛋白組方案,翻譯后修飾GPS導航,助力您的文章發表,歡迎垂詢!


系統生物學研究內容主要包括“基因-蛋白-代謝-表型”等多個層次,越來越多研究表明,多組學已成為生命科學和醫學研究的重要工具。“基因組反應了可能發生的變化,蛋白組和代謝組反應了正在或者已經發生的變化”,邁維代謝持續創新質譜技術,創新性的開發出了廣泛靶向代謝組檢測技術 TM,建立了行業領先的植物代謝數據庫和醫學代謝物數據庫 MWDB,真正實現了“高通量、超靈敏、廣覆蓋”盡可能多的檢測樣本中所有的小分子化合物。

與此同時,邁維代謝和賽默飛達成戰略合作,詳情請點擊:拓展組學研究的邊界 | 邁維代謝攜手賽默飛共建戰略合作實驗室。在引進新一代質譜平臺 Orbitrap?Astral?并同步配置 Orbitrap Exploris 120 質譜儀后,雙方進一步深入技術聯合開發,攜手努力打造為世界領先的創新蛋白質組和代謝組研發中心。加快和深化對拓展蛋白質組學及多組學領域的探究,更好地服務于生命科學和醫學健康研究領域,助力精準醫學高質量發展!








評論